|
||||||||||||
            |

Szexuális szelekció
| A lényeg: |
A természetes és szexuális
szelekciót sokszor külön kategóriaként tekintették,
hiszen sok esetben külön jellegekre hat, vagy gyakran ellentétes
az irányuk, de lényegében a szexuális
szelekció a természetes szelekció azon része,
amikor a forrás, melyért az egyedek között a versengés
zajlik, az a potenciális szexuális partner, a pár. Ugyanúgy,
mint egyéb környezeti vagy biológiai tényezõk,
melyek szükségesek a túléléshez, és
ezzel a gének hatékony továbbadásához,
szexuális szaporodásnál a partner is szükséges
az utódok létrehozásához. A
szexuális szelekció így lényegében az a
szelekció, ami azokra a jellegekre irányul, melyek a párért
folyó versengésben valamilyen módon esélyesebbé
teszik a versenyzõket.
Két formáját különböztetik meg, az úgynevezett
intraszexuális szelekciót, ami a versengõ nemen
belül folyik az elérhetõ párok monopolizálásáért
(és az az ivar, amelyikért a versengés folyik, nem szól
bele a verseny kimenetelébe), és az interszexuális,
vagy nemek közötti szelekciót, ami
lényegében az egyik nem preferenciája bizonyos, a másik
nemen belüli gaméták és hordozóik felé
(itt a versengõ ivar lényegében a preferenciáknak
igyekszik megfelelni). Az "interszexuális" kifejezés ez esetben
nem a legszerencsésebb, hiszen lényegében ez is a nemeken
belül folyó versengés az elérhetõ partnerekért,
a választók is nemtársaik esélyeit csökkentik
(pl. egy jó minõségû egyed monopolizálásával).
A szexuális szelekció elsõdleges
oka az anizogámia. Ha már kialakult a szexuális
szaporodás és az anizogámia (lásd az elõzõ
fejezeteket), az jó oka és alanya a szexuális konfliktusnak.
Az anizogámia miatt a két nem primer szaporodási befektetése
aszimmetrikus (vagyis az ivarsejtek szintjén a hímek általában
sokkal kevesebbet fektetnek egy darab utódba), és ez az eredeti
aszimmetria tovább erõsödhet a szülõi gondoskodás,
vagy egyéb befektetések nemek közti különbségével.
Így a potenciális utódszám is jelentõsen
különbözik a nemeknél, és emiatt az egyik nem
limitálja a másik szaporodási sikerét. Ez általában úgy néz ki, hogy a
populációban jóval kevesebb megtermékenyíthetõ
petesejt van, mint amennyit az ivarsejtek képesek lennének fertilizálni
(1:1 ivararánynál az ivarsejtek aránya 1:nagyon sok a
hímek javára)
Vázlatosan:
Nem
befektetõ nem (általában a hím): |
Befektetõ nem
(általában a nõstény): |
Kis szülõi allokáció
egy utódba |
Nagy szülõi allokáció
egy utódba |
Nagy szaporodási potenciál |
Kis szaporodási potenciál |
Szaporodási siker
növelése a párok számának növelésével |
Szaporodási siker
növelése a párok minõségének
növelésével |
Párért való versengés
nagy |
Párért való versengés
kicsi |
Szaporodási siker varianciája
nagy |
Szaporodási siker varianciája
kicsi |
Limitáció kicsi |
Limitáció nagy |
Kompetitív nem |
Választós nem |
A versengõ nemen belül a szaporodási siker varianciája nagy lesz, a rátermett egyedek sok párt tudnak kisajátítani, míg a rosszabb fitnessûek alig, vagy egyáltalán nem jutnak párhoz. Ezzel szemben a limitáló ivar szaporodási sikere sokkal kiegyenlítettebb, egy rosszabb minõségû egyed is nagy valószínûséggel tud szaporodni, hiszen a versengõ egyedek (illetve gaméták) feleslegben vannak hozzájuk (illetve gamétáik számához) képest.
A kompetitív nemen belül a versengés mértéke megnõ, ha a párok elérhetõsége csökken, mert
1. az ivararány, vagy az
operacionális
ivararány
(operational sex ratio, OSR: az adott pillanatban, a
populációban
elérhetõ szaporodóképes hímek
és
nõstények aránya) lecsökken
2. az élõlények
szemelpárok (csak
egyszer
szaporodnak életükben), és a fitnessük csak ez
egyetlen
szaporodási eseménytõl függ (mindent vagy
semmit)
3. az ivarzó egyedeket a
hímek nagyban tudják
kontrollálni, mert pl. a nõstények nem egyszerre,
nagy tömegben, hanem külön ivarzanak,
lehetõvé téve
a monopolizálásukat.
A kompetíció ökológiailag tekintett formái
alapvetõen kétfélék lehetnek. Az interferenciális
(kiszorítós) kompetencia feltételezi az egyedek közvetlen
kölcsönhatását, fizikai jelenlétüket,
míg az exploitációs (kizsákmányoló)
kompetenciához nem szükséges mindkét versenytárs
jelenléte, az egyik egyed azzal is csökkentheti a másik
forrásait, hogy eleszi, elszedi, elszívja elõle, és
az a másik számára már nem lesz elérhetõ.
Ezek analógiájának tekinthetõ a szexuális
vetélkedés kétféle formája, az intraszexuális
(küzdő, kitartásos vagy tülekedő, lásd később)
versengési formák, amik direkt módon, és az interszexuális
párválasztás (mate choice), ami indirekt befolyásolja
az egyedek párhoz jutását.
Fordított ivari szerepek
Bizonyos fajoknál az általános mintázat megfordul, a hímek lesznek az inkább befektetõ nem. Ilyenkor a õk jelentik a limitáló tényezõt, hiszen a kevesebbet befektetõ nõk több utódot tudnának létrehozni, mint amennyire a hímek, a versengés a hímekért folyik. Ez akkor fordul általában elõ, ha egyéb tényezõk beleszólnak a gametikus befektetésbe, és elfedik az anizogámia eredeti hatását.
|
A mormon tücsöknél a hímek egy nagyon nagy, és energiában gazdag spermatékát választanak ki, ezt a nõstény megeszi. Ebben vannak a spermiumok is, plusz rengeteg tápláló fehérje, amire a nõsténynek szüksége van a tojástermeléshez. A hím csak egyetlen spermatékát képes termelni, és így a nõstények számára nagyobb limitáló tényezõ lesz, mint a sok tojást termelõ nõstény. Ezért a nõk versengenek és harcolnak a hímekért. A Syngnathus tûhalnál a hímek nagyobb szülõi aktivitást fejtenek ki, mert õk hordozzák a kicsiket a költõtasakukban. Ez idő alatt nem tudnak más nőstényeknek udvarolni, míg az anyai terhek alól felszabadult lányok más hímek után nézhetnek. A nõstények nagyok és színesek, és harcolnak a hímekért, a kicsi és fakó hím választ közülük (ált. a nagyobbat és színesebbet). A poliandriás
(lásd később) jasszána nõsténye nagyobb és
agresszívebb, mint a hímek. A nõstények
védik a territóriumokat, és a rajtuk kotló
hímeket.
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
A párért való versengés formái
1. Intraszexuális
mechanizmusok
Ilyen jellegű
kompetícióban a versengő nem
(ált. a
hímek) egyedei a másik nem aktív
részvétele nélkül próbálnak
minél több szaporodási sikert elérni.
Prekopulációsan
1. Küzdõ versengés
(contest
competition)
Ez lényegében a
versengõ nemen belüli
direkt harcot jelöli, amikor az erõsebb, ügyesebb,
agresszívabb egyed elüldözi a nõstény
közelébõl a gyengébb
vetélytérsa(ka)t. Ilyen vetélkedés
esetén szexuális szelekció hat a nagy
testméret, a különbözõ fegyverek az
ügyesség és bizonyos riasztó jelzések
kifejlõdésére.
2. Kitartásos
rivalizálás (endurance
rivalry)
Ekkor a versengésben az az egyed
lesz gyõztes, amely a
legtovább képes reproduktívan aktív
maradni. Ekkor a szelekció kedvez a jó
kondíció
és kitartás képességét
megnövelõ jellegeknek. Ez a természetes
szelekcióban is lehet
elõnyös, de néha ellen is hathatnak.
3. Tülekedõ
versengés (scramble
competition)
Az lesz itt a gyõztes, aki a legkorábban képes felfedezni
és megtermékenyíteni a nõstényeket. Elõnyös
a gyorsaság, a korai szexuális érés,
a fejlett érzékszervek és térbeli memória
fejlettsége.
{kind=link}
Posztkopulációsan
1. Indukált abortusz,
infanticídium
(gyerekölés)
Ez a viselkedésforma a vetélytárs hímek utódainak
elpusztításával közvetlenül csökkentheti
azok szaporodási sikerét és a nõstényt
újra fogékonnyá tehetik a szaporodásra az infanticid
egyeddel (pl. oroszlánok és langúrok).
2. Spermakompetíció
Ez a kevésbé szembetûnõ versengési forma
lényegében a nõstény ivari traktusában
játszódik le, és a résztvevõk maguk a spermiumok.
Ha több hímmel is pározhat a nõstény, az
apaság valószínûségét sok mechanizmussal
lehet növelni. A jó spermakompetitornak pl. sok spermiuma és
nagy heréi lehetnek, elõnyös lehet a többszöri
párzás, a nõstény elválasztása és
õrzése. A kopulációs mirigyek és szervek
is lehetnek a szex. szelekció alanyai. A mai elgondolások szerint
a hím genitália lényegében belsõ udvarló
szerkezet, és sokféleségüket nem elsõsorban
a faji elkülönülés elõsegítése
(kulcs-zár modell, természetes szelekció alatt), hanem
a szexuális szelekció hajtotta. (Sok rovarnál a hím
ivarszervek igen, de a nõik nem mutatnak erõs faji különbségeket,
pedig a nõstény számára a más fajjal történõ
párzás sokkal költségesebb, tehát a kulcs-zár
valószínûleg nem jó magyarázat).
| A párõrzés
hatékony fegyver lehet a kompetens hímek kiszorításában.
Gammarus rákok párzás
elõtt és után sokáig szorosan tartják
életük párját, megakadályozva a többi
hím közeledését.
Számos furfangos mechanizmus fejlõdött ki a vetélytársak esélyének csökkentésére. A feketeszárnyú szitakötõ péniszén (és számos más rovarfajén is) olyan horgok találhatók, melyek lehetõvé teszik az õt megelõzõ hímek spermájának kiseprését a nõi nemi utakból. Linyphia
pók párzás után a nõt olyan
feromonnal keni be, amirõl a többi hím hímnek
hiszi, és nem környékezi. |
{kind=link}
{kind=link}

2. Interszexuális mechanizmusok
Ez a versengési forma sokkal kevésbé "direkt", a nem
versengő ivar aktívan alakíthatja a verseny kimenetelét
a "nyertes" valamiféle kiválasztásával
Prekopulációsan
1. Hölgyválasz (mate choice)
Itt az az egyed lesz sikeres, amelyik az ellenkezõ nemnek (ált.
nőstény) tetszõ jellegeket birtokol, a versengés
itt a preferenciáért folyik.
Posztkopulációsan
1. Rejtett hölgyválasz (cryptic
female choice)
A spermakompetíció
folyamataiba is beleszólhat
a
választó nőstény. A mai szemlélet szerint a
nõstény
nem csupán a spermakompetíció passzív
helyszíne, hanem ezen a szinten is aktívan
kontrollálja
verseny kimenetelét és az utódok
apaságát,
és a hölgyválasz ezen a szinten is
érvényesülhet.
| Madaraknál
megfigyelték, hogy párzás után a
nõstény kipréseli
kloákájából az ondót.
Sárga trágyalégynél a nõ aktívan eltávolíthatja bizonyos hímek spermiumát más spermiumok javára az erre hivatott receptákulumból (nem a versenytárs hím csinálja, pénisze el sem ér odáig párzáskor). Diplosoma listerianum
szivacsnál (és nagyon sok
növénynél) bizonyos spermiumok
kívánatosabbak, mások megemésztõdnek
|
{kind=link}
A nõstények és a többszörös
párzás
A többszörös
párzás sokféle
káros hatással járhat a nõstény
számára (lásd szex.konfliktus), mégis
sok fajnál
megfigyelték, hogy nõstény aktívan keresi
a sok hímmel való kopulációt.
Kétféle magyarázat
lehetséges:
1. Jó genetikai állománnyal rendelkezõ
(superior) hím spermáját keresi. Ha a hím
kvalitását nem jelzi semmi, ezalapján nem tud
dönteni, ekkor a sperimumos szintjén dõl el a
jóság (spermakompetícióval vagy rejtett
hölgyválasszal).
2. Genetikailag kompatibilis hímet keres, nem valami általánosan "jó" egyedet.
|
Cordilochermes scorpioides álskorpiónál a többszören párzott nõstények utódjai jobban túléltek, ez nem korrelált hímek minõségével, valószínûleg génkompatibilitás van a háttérben. Egérnél MHC heterozigóták jobb túlélést mutattak, jó a másfélével párosodni, ezt szag alapján meg is képesek találni. Fürge gyík többszörös párzása csökkenti a beltenyésztést és a deformált utódok arányát |
{kind=link}
{kind=link}

A többszörös párzásnak
vannak igen komoly hátrányai és költségei
is a nõstények számára, ezekrõl késõbb
lesz szó.
Miért választanak a nõk bizonyos hímeket, és
miért mellõznek másokat? A nõsténynek
primer kontrollja van az utódnál, eldöntheti, hogy melyik
utódba (petébe) mennyit invesztál, de válogathat
a potenciális apák közül is, sõt még
az egyes hímek spermái közül is. A nõk reproduktív
döntését persze a hímek befolyásolhatják
források szolgáltatásával, udvarlási
viselkedéssel, esetleg erõszakkal vagy gyakori párzással.
Direkt haszon
választása
A legegyértelmûbb dolog
az, ha bizonyos hímek
nagyobb
anyagi forrást nyújtanak a nõnek, mint
mások. Ilyen anyagi haszon lehet a nõstény
(és így
az utód) számára hasznos
táplálék vagy ásványi anyag,
érdemes az ezt (jobban) nyújtó hímet
választani.
| Bittacus csüngõlegyek hímje
párzáskor rovarzsákmányt ad a nõsténynek.
A párzás hossza az adott préda méretétõl
függ (meddig csócsálja a nõ). Ha kicsi a zsákmány,
hamar végez vele és kicsi az esélye a spermiumok
bejutásának. Legalább húszperces zsákmány
kell.
Gluphisia lepke hímje párzás
elõtt sok vizet iszik pocsolyából, melybõl
kivonja a ként, és a spermatofórájába
építi. A nõstény ezt megeszi, és
így a kén az utódba jut, segítve fejlõdését
a kénhiányos tápnövényen. |
{kind=link}
{kind=link}
Indirekt haszon
választása
Más fajoknál nem tűnik
ilyen egyértelműnek a
nőstények választása. A lek
poligíniás állatoknál (lásd
később) pl. a nőstények a hímektől
semmiféle befektetést nem kapnak, kizárólag
a spermával "szállnak be" az utódokba. Ekkor
más, bonyolultabbnak tűnő hasznok
preferálását sejtik a nőstények
választása mögött, és ezek
magyarázására sok teória született.
Íme a legfontosabbak:
1. Elfutó (runaway) szelekció
1903-ban Morgan éles kritikával illette Darwin 1871-es, a szexuális szelekciót tárgyaló könyvét, mivel a nõi preferencia jelenségérõl nem tesz említést. Fisher ezt a kritikát olvasva gondolta ki a teóriát. A modell feltételezi, hogy a hím jellegben is van variancia a populáción belül (mondjuk, vannak rövidebb és hosszabb farkú egyedek), és a jelleg iránt való nõi preferenciában is (némely lánynak a rövidebb tetszik, másoknak a hosszabb), valamint a jelleg is és a preferencia is öröklõdik.
Mechanizmusa: Adott egy tulajdonság (pl. hosszú farok), ami kezdetben együtt jár valamiféle (akár enyhe) fitness-elõnnyel. Ekkor érdemes a populációból az ilyen srácokat választani, mivel így az e hímektõl származó gyerekek is rátermettebbek lesznek. A populációban elterjedhet a hosszú farok preferálása. Viszont a hímek versengésében szelekció fog hatni a hosszabb farok kifejlõdésére, mivel ezek könnyebben találnak párt, nagyobb a szaporodási sikerük. Mivel érdemes olyan fiat szülni, amelyiknek a nagy preferáltság miatt nagy a szaporodási sikere, (sexy son), továbbra is érdemes az egyre hosszabb farkúakat választani, mivel akkor a fiaknak is nagy sikerük lesz. Ebben a fázisban már nem játszik szerepet, hogy a jelleggel ténylegesen jár-e együtt a hímnek valamilyen jó tulajdonsága, mint kezdetben. Itt a jó tulajdonság a kelendõség. Végeredményben ez egy pozitív visszacsatolású, önerõsítõ, elfutó folyamatot eredményez. Egyre nagyobb farkú hímeket a lányok egyre jobban szeretik, ezért egyre jobban megéri hosszú farkú hímnek és hosszú farkat kedvelõ nõnek lenni. Persze a hosszú farok más szelekciós tényezõk által korlátozva van, nem nõhet a végtelenségig. De ha megfelelõ a preferencia, a jelleg akkor is elterjedhet ha viselõjének valamennyire hátrányos (pl. növeli a predáció veszélyét). A lényeg, hogy a fiak szaporodásisiker-elõnye miatt érdemes a preferált tulajdonsággal bíró hímeket választani.
|
A popslágerlisták hasonló
pozitív visszacsatolással mûködve
befolyásolják a fogyasztók döntéseit.
Minél inkább kedvelt egy sláger (a lista
élén van), annál inkább kedvelt lesz
mégjobban, mert sokan éppen emiatt veszik meg. Nem is
kell a számnak jónak lenni,
elég ha valahogy a listára kerül, utána
már
beindul az önerõsítõ mechanizmus. |
2. Indikátor (jó
gén) mechanizmusok
Ezt a teóriát Wallace
már 1889-ben
felvetette, és ezt is Fisher öntötte modern
formába (bár az elõzõ modellel
kacsolják össze a nevét fõleg). Ez a modell
olyan jellegeket feltételez, melyek a viselõjének
viabilitását, fitnessét tükrözve
fejezõdik ki, valamint a másodlagos
jelleg és a fitness-tulajdonság kapcsoltan
öröklõdik. Ha az ezzel járó
fitness-elõny örökíthetõ, akkor
érdemes az ilyen hímet választani, mivel
rátermettebb utódokat eredményez. Tehát az utódokba jutó
elõnyös gének miatt érdemes bizonyos
hímeket választani.
| A slágerlista-analógiát
használva, itt tényleg a jó számok
kerülnek a top10-be. |
Zahavi ('75) értelmezése szerint a hendikep a lényeg a dologban. Amely hím sikeresen dacol a hosszú, színes, stb. feltûnõ jellegek megnövekedett költségével, az elég rátermett, és a nõk emiatt preferálják.
Egyes értelmezések
szerint az életkor egy jó általános
indikátora a fitnessnek, mivel az a hím, amely idősebb
kort ért meg, bizonyította
rátermettségét (jó génjeit) a
paraziták, predátorok, stb. ellenében, és
érdemes választani. Viszont a korral együtt
csökkenhet a fertilitás és az ivarsejtekben
bekövetkezett káros mutációk esélye is
nagyobb.
Hamilton és Zuk ('82)
modellje szerint
(parazita-mediált szexuális szelekció) a
paraziták okozhatnak idõben gyorsan
váltakozó szelekciós nyomást, és a
másodlagos jelleg sok esetben jó immunitást
okozó génekkel járnak együtt. A
feltûnõbb jelleg jobb
ellenállóképességet mutat, érdemes
választani az utódokba
örökítõdõ jobb immunitás miatt.
Természetesen a fisheri és indikátor
mechanizmusok nem egymást kizárók, együtt is hathatnak.
Az elõzõ két modell nem direkt hasznot feltételezett,
hanem valami késõbb jelentkezõ genetikai elõnyt.
Lehetnek azonban közvetlen nyereségei is a nõstény
számára az adott hím preferálásának,
ez esetben lényegében fenotípusos varianciát választ
a nõ (pl. parazitamentes hímet, ami nem fertõzi meg õt
és gyerekeit).
3. Párválasztás
és genetikai kompatibilitás
A jó gén elméletnek sok bizonyítéka van,
fõleg vizuális szignáloknál, ezek megfeletlethetõk
fitness- haszonnal járó jellegekkel (pl. pirosabb taraj-jobb
rezisztencia). Sok újabb eredmény szerint viszont a genetikai
kompatibilitáson alapuló hím-preferencia is gyakori.
Ezt nehezebb szignálokkal elképzelni (kinek milyen piros mit
jelent?). Az ismert bizonyítékok a pár kiválasztásában
fõleg olfaktorikus csatornákon történõ információ-szerzést
sugallnak (hal, ember, rágcsálók, MHC). Ahol ez az út
sem járható, ott a nõstény többszörös
párzással biztosíthatja a kompatibilitást (valamelyik
majd csak passzol). Itt az utódok jó heterozigóciája
miatt érdemes bizonyos hímeket választani.
A jó gén- mechaizmus és a kompatibilitás szimultán
mûködhet, pl szociális párnál jó gént,
EPC-párnál kompatibilitást kereshet a nostény.
| Itt mindenkinek
saját slágerlistája van, mindenkinek más
a jó. |
Az ivari dimorfizmus fõbb formái
Az egyedek reproduktív
sikerüket sokféle
módon növelhetik, és az ökológiai vagy
élettani viszonyok is különbözõek az egyes
taxonokban,
így számos és sokféle tulajdonság
állhat szexuális szelekció alatt. (Pl. az
elsõsorban
látásukat használó állatoknál
vizuális jellegek, a fejlett szaglásúaknál
a kémiai szignálok, stb.)
1. Testméret
A testméret esetében
természetes és
szexuális szelekciós hatások egyaránt
szerepet játszhatnak, a nagy test a túlélés
számos aspektusában elõnyös lehet.
Nagy hím testméret adaptív akkor, ha küzdõ versengés van a nõstényekért, vagy ha territóriumot kell védeni, hasznos lehet a kitartásos rivalizálásban és a spermakompetícióban (párõrzés, elkülönítés hatékonyabb)
Kis hím sikeres lehet, ha a küzdelem háromdimenziós térben játszódik (víz, levegõ), mert ez gyorsasággal, ügyességgel párosulhat, vagy tülekedõ kompetíció esetén, mert itt is gyorsabb lehet a kicsi.
Nõsténynél a nagy testméret erõs természetes szelekciós hatás alatt is áll, a nagy nõsténynek jó a reproduktív sikere, azon egyszerû oknál fogva, hogy sok és nagy petét tud termelni, vagy jó szülõi gondoskodást tud nyújtani (ennek nincs köze a párért való versengéshez). De a szex. szelekcióban is kedvezményezett lehet a nagy nõstény, ha mondjuk õ véd territóriumot vagy verseng a hímekért (fordított ivari szerepnél)
Kis nõstény sikeres lehet, ha korai érés elõnyt jelent (lenyúlja a többiek elõl a legjobb hímeket vagy forrásokat), vagy ha hirtelen kell energiát mobilizálni a tartalékokból.
Legtöbb állatban a nõstény nagyobb, mint a hím. Általános jelenség, hogy ahol a hím nagyobb, ott annak oka a vetélkedés jellege, ami küzdõ versengés vagy territorialitás (pl. emlõs madár, bogár, hüllõ).
Törpe hímek sok
fajnál elõfordulhatnak,
itt
a nõstények sokkal nagyobbak (pókoknál vagy
mélytengeri halaknál, sokféle
endoparazitánál). Általában ott van ilyen
nagy méretkülönbség, ahol szesszilis és
nehezen megtalálható a nõstény, ekkor a kis
hímméret kedvezhet a migrációs
képességnek (pl. pókfonálon) és
nincs lehetõség a nagy méretet
preferáló küzdelemre. Mélytengeri
halaknál a hím, ha rátalált a nõre,
parazitaként rá is akaszkodhat, ezzel biztosítva a
szaporodást (nehéz új lányt találni,
és ha már megtalálta, így a legjobb vele
maradni (és esetleg megvárni, míg ivarérett
lesz).
2. Fegyverek
Különbözõ
agancsok, karmok, agyarak,
számos állatcsoportban kifejlõdtek. Sok
hipotézis született, hogy miért (predátor
ellen, más hímek
ellen, indikátor mechanizmusokban játszik szerepet).
Mivel csak kevés fajnál van a nõstényeknek fegyvere,
így valószínû, hogy a hím-hím vetélkedés
hajtotta az evolúciójukat. Ahol nõsténynél
is van (rénszarvas, néhány antilop),
ott szerepe lehet a ragadozók ellen való védelemnek.
A hölgyválaszban is lehet szerepe (tükrözheti a viselõje
kondícióját, korát védelmezõ képességét).
Emlõsöknél sokféle
fegyver lehet, a testmérettel változik
a verekedési stratégia. A kis testûek inkább hemperegve,
a földön, fogakkal vagy karmokkal harcolnak, míg a testmérettel
a földre kerülés miatti sérülés veszélye
nõ, hasznosabb állva alkalmazható fegyvereket fejleszteni.
Ha olyan fegyvert akarnak használni, amely nem sebesíti meg
a vetélytársat, de az egész test erejét bele lehet
vinni, úgy célszerû a fejen lévõ agancsokat,
tülköket kifejleszteni. A párosujjú patások
evolúciója jól illusztrálja a folyamatot. Az õsi,
kistetstû fajok, mint a dik-dik antilop vagy a pézsmaszarvas szarvatlanok, és hemperegve,
fõleg a szemfogaikat használva küzdenek. A nagyobbaknál
megjelennek a szarvak, és egyre nagyobbak lesznek a testmérettel.
A küzdés ezeknál már állóhelyzetben,
"karambolozva" történik.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Madarak esetében a lábon és szárnyon lévõ sarkantyúk (tyúkfélék) és karmok (pl. pingvinfélék) lehetnek ilyen szerepûek. Rovaroknál, fõleg a bogaraknál feltûnõ a fegyverek dimorfizmusa (szarvasbogár, herkulesbogár, szarvaslegyek). |
{kind=link}
{kind=link}
3. Vizuális szignálok
Feltûnõ színek és formák rengeteg féle
állatban kifejlõdtek, ezek nagy része pont a szexuális
szelekció miatt, amire bizonyíték, hogy sok ilyenféle
jelleg csak a nemi érettséggel jelentkezik, vagy csak a hímek
rendelkeznek vele, esetleg csak a párnak és rivális felé
jeleznek vele, vagy csak a szaporodási szezonban vannak jelen. Az ilyen
szignálok "szerelemre és háborúra" egyaránt
jók lehetnek, szolgálhatnak a versengésben és
a hölgyválaszban is.
Sok feltûnõ jelzés szolgálhat aposzematizmust is (ragadozó-elrettentés), ekkor általában mindkét nemnél jelentkezik.
4. Akusztikus jellegek
Ezek a jellegek a leginkább
szexuálisan
szelektáltak, sok szignál csak a versengés miatt
jött létre. Persze más, a szaporodási sikert
szintén növelõ hangjelzések nem
szexuális szelekció alatt állnak (pl.
vészkiáltás). Fõleg rovaroknál,
békáknál és madarakban jellemzõ,
de halak és emlõsök is élhetnek vele.
Szintén szerepelhet contest cometition és female choice
mechanizmusokban is.
| Egy másféle
szignál, amikor egy domináns bika háremébe
tartozó nõstény jelzi a háremúrnak
ha egy szubdomináns másik hím akar vele párzani,
és az így elkergeti a trónkövetelõt
(pl. elefánt, elefántfóka) |
{kind=link}
5. Kémiai szignálok
Szintén elterjedtek, és sok esetben állnak szexuális
szelekció alatt. Használhatják párok vonzásra
(rovarferomonok), riválisok elrettentésére (pl. halak),
szexuális stimulálásra (gõték). Emlõsöknél,
mint alapvetõen olfaktorikus állatoknál, nagy szerepet
kap.
A másodlagos jellegek kényszerei és korlátai. A nemi jellegek elvesztése
A szexuális szelekció alatt álló jellegek sok esetben kerülnek szembe egyéb szelekciós erõkkel. A jelleg, mely nagyban segíti a viselõjét a párért folyó versenyben, hátrány lehet más vonatkozásban. Sokféle formában és fejlõdési stádiumban jelentkezhet ez a konfliktus. Ezenkívül a szexuális szelekció is lehet önszabályozó, a nemi jelleg bizonyos esetben növelheti, más tekintetben csökkentheti a szaporodási sikert.
1. A predációs nyomás jelentõsen megnõhet, ha a nemi szignál a ragadozó számára is feltûnõbbé teszi a kibocsájtót, pedig az csak a pár vagy a rivális számára akart feltûnõ lenni.
| Hóbagoly-köpetek
tartalmát vizsgálva azt figyelték meg, hogy azokban
több a hím barna lemming maradvány, mint a nõstény.
Vándorsólyomnál ugyanez
volt a helyzet cankó
zsákmányállattal. Persze ez nem bizonyítja
közvetlenül, hogy a hímek jobban
feltûnnének a ragadozóknak, lehet, hogy a
hímek nagyobb aktivitása és szem elé
kerülése az oka az eltolódott
préda-ivararánynak. |
{kind=link}
| Tüskés pikóknál a feltûnõ
piroshasú hímek gyakoribb zsákmányai a ragadozóhalaknak,
mint a fakó nõstények. Trinidadban olyan guppipopulációknál, ahol a perdációs
veszély nagyobb, ott a hímek kevésbé viselnek
feltûnõ színeket, mint a gondtalanabb életû
populációknál.
A tûzbogaraknál a nemek párzási
idõben fényjelekkel jelzik egymásnak hajlandóságukat.
A Photinus fajok párvonzó fényjeleit
utánozzák a ragadozó Photuris tûzlégyfajok, zsákmányolásra
használva a Photinus hím gerjedelmét. A Photuris
fajok hímjei viszont a Photinus jelet utánozva, magukat
prédának álcázva csalják “szobára”
az éhes nõstény fajtársakat. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
|
Rövidfarkú tücsök esetében a ciripelõ hímek több estben lettek gémek prédái, mint csendes, szatellit társaik. Mezei tücsök ciripelése is jobban vonz egy parazita légyfajt (persze a nõstényt is), mint a csendesek.A tungara béka éneklõ hímjei is gyakrabban végzik a denevérek gyomrában, mint a csendes szatellit egyedek. |
{kind=link}
{kind=link}
{kind=link}
| Pókok
hálójában többet találni bizonyos
repülõ rovarok hímjeibõl, mint a
lányokból.
Valószínûleg a hímek aktívabban
és
többet repülnek párt keresve, a
nõstények
otthonülõsebbek. |
2. Fiziológiai kényszerek
A display-mutatványok, nászrepülés- és úszás,
a csatározások, valamint a hosszú párosodási
szezon igen energiaigényes aktivitások, így ezek a túlélést
nagyban csökkenthetik.
3. Genetikai kényszerek
Sok nemi jelleg nem ivari kromoszómán található,
de csak az egyik nemben fejezõdik ki hormonhatásra. Azok a jellegek,
melyek hasznosak egyik nem reproduktív sikerének növelésében,
károsak lehetnek a másik nemnél, így (ha autoszómán
örökítõdnek) ez a hátrány határt
szabhat evolúciójuknak. Szelekció hat a csak az egyik
nemnek elõnyös másodlagos jellegek génjeinek nemi
kromoszómákra kerülésére, mert azok így
klonálisan, csak az egyik ivarba adódnak át, kikerülve
ezzel a problémát.
4. Érzékszervi eltolódás
(sensory bias)
Lehet káros a preferencia olyan estben, amikor egy adott faj egyede
olyan jelleget preferál, ami csak egy másik fajnál található
meg, így félrepárzás következhet be, ami
steril vagy életképtelen hibrideket eredményezhet.
A szexuális szelekcióval foglalkotó vizsgálatok általában "egyoldalúak", a jellegek kialakulására és fenntartására irányuló erõk szerepét állítják a középpontba. Filogenetikai vizsgálatok azonban rávilágítottak egy tényre, amely eddig elkerülte a kutatók figyelmét. A rokon fajok leszármazási vizsgálataiban számos esetben figyelték meg a szexuális jelleg vagy a jellegre irányuló preferencia elvesztését, és ez széles körben elterjedt evolúciós trendnek bizonyult, a legkülönbözõbb taxonokban találtak rá példát.
A jelenség magyarázata nem egyértelmû, sokféle mechanizmust sejtenek mögötte. Általában olyan esetben feltételezik a szexuális szelekció miatt kialakult jegyek elvesztését, amikor egyéb szelekciós erõ vagy a genetikai sodródás hatása erõsebbnek bizonyul, mint a szexuális szelekció.
Az okok között szerepelhetnek:
1. Környezeti
tényezõk:
Predációs nyomás
megnövekedése
ellensúlyozhatja a szexuálisan szelektált jelleg
elõnyeit, ekkor érdemes lehet azokat "elfelejteni".
| Guppi
hímek ragadozóveszélyes helyen halványabbak,
mint gondtalanabb populációban élõ társaik.
|
{kind=link}
{kind=link}
Jelzés hatékonyságának csökkenése is lehet elõnytelen bizonyos szignálok esetében.
| Tüskés pikó hímek sötét vízben élõ populációiban a piros hasfolt eltûnt |
{kind=link}
Táplálékellátottság is befolyásoló tényezõ lehet.
| Carpodacus mexicanus pinty bizonyos populációiban
a karotinoid-tartalmú táplálék ritkasága
miatt a karotinoid tartalmú színes tollazat eltûnt.
|
{kind=link}
2. Szociális folyamatok
|
Az örvös és kormos légykapó nõstényei olyan populációkban, ahol a két faj nem található meg együtt, egyaránt a feltûnõ fekete-fehér mintázatú hímeket preferálják. Ahol viszont átfed a két faj, ott a kormos légykapó lányok hímjeiket minél kevésbé feltûnõ színnel szeretik, valószínûleg a félrepárzást elkerülendõ (ez szelekciós erõ a hímek színpompájának csökkenésére). Házi verébeknél a nõstények bizonyos populációkban a kis begyfoltú, kevésbé agresszív és jó utódgondozó hímeket szeretik, máshol a nagyfoltú, verekedõs, jó fészkelõterületet szerezni képes hímeket. Poephila acuticauda pintyfajnál a hím
feltûnõ színezetének elvesztése
nõstény-utánzást, és ezzel a hím-hím
agresszív interakciók csökkentését
szolgálhatja. |
{kind=link}
{kind=link}
{kind=link}
3. Véletlen tényezõk.
Ha a szexuális szelekció gyenge, úgy (fõleg kis
populációkban) a genetikai sodródás könnyen
az adott jelleg elvesztéséhez vezethet.
A szaporodásisiker-maximálás nemek közti különbsége olyan folyamatokhoz vezethet, mely az egyik nem sikeressége érdekében káros hatással van (nincs tekintettel) a másik nemre, vagyis az adott jelleg elõnyös a versengõ nemnek, de valamilyen módon káros a másik ivarra. Az érdekek konfliktusa alakíthatja a szexuális szelekció mûködését és jellegét. A hímek és nõstények érdekei eltérõek, így a párzás és utódnevelés során a felek számos konfliktussal szembesülnek. Ezeket feloszthatjuk prezigotikus és poszt-zigotikus (szülõ-szülõ) szexuális konfliktusra (errõl késõbb).
Prezigotikus szexuális konfliktus
Egy sikeres hím, mely legyõzi vagy "túljelzi" versenytársait,
sok nõstény monopolizálásával sikeresen
növelheti szaporodási sikerét, akkor is, ha a párzott
nõstények számára ez valamilyen költséget
jelent, és reproduktív potenciáljuk csökkenésével
jár. Minél nagyobb a szaporodási siker varianciája,
annál intenzívebb a párért való versengés,
és annál inkább valószínû, hogy
a nõstény számára esetlegesen káros
jellegek megjelennek. Sok vizsgálatból kiderült, hogy
számos, a hímek versenyében elõnyös jelleg
vagy viselkedés csökkentheti a nõstény túlélését
és fekunditását, vagy egyéb módon hátrányos
a számukra.
Gyakori a nõstények megtévesztése, vagy a
nem-reproduktív jelzések kihasználása
|
Emphis legyek hímjei selyemgubóba csomagolt nászajándék zsákmányt adnak a nõsténynek párzáskor. Sok esetben a cseles hímek csak üres gubót adnak ajándékba, így megúszva a költséges (és a nõsténynek nagy indeirekt hasznot jelentõ) zsákmányolást. Corynopoma pontylazacoknál a hímek úszóin olyan függelékek fejlõdtek ki, melyek a zsákmányt imitálnak, és ezekkel a hím az éhes nõstény figyelmét könnyen fel tudja kelteni. A Photinus és Photuris
tûzbogarak példáját lásd fentebb |
{kind=link}
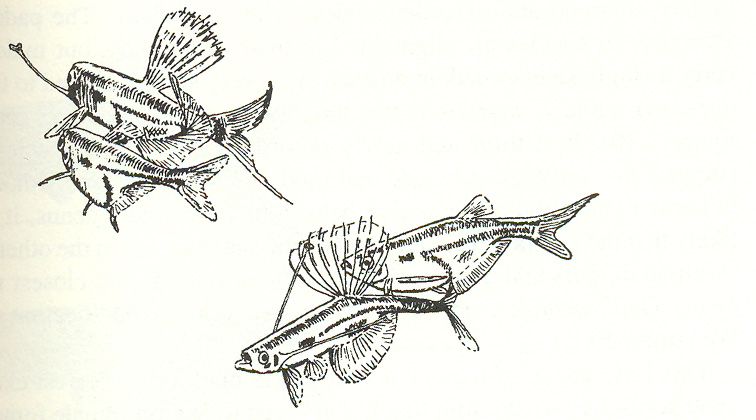{kind=link}
A más hímektõl származó utódok elpusztítása (infanticídium) nagymértékben növeli a hím esélyeit (a nõstény abbahyja a nevelést, és újra fogékony lesz), míg a nõstényeknek hatalmas befektetés elvesztése.
| Oroszlánok, Langúrok, de pl. Stegodyphus pókok hímjeinél jól ismert. Fordított ivari szerepeknél a Jasszánák nõstényeinél is van ilyen. |
Az erõszakos párõrzés, a nõstényért való dulakodás vagy az elnyújtott, többszöri vagy kényszerített kopulációk (nemi erõszak) sok esetben lehetnek károsak a nõsténynek, mivel korlátozzák a hölgyválaszos döntést, valamint akár csökkenthetik is túlélését és fekunditását.
| Cimex lectularius ágyi
poloskák hímjei a nõstény testfalát
átfúrva közvetlenül a testüregbe ürítik
a spermiumjaikat, csökkentve ezáltal a nõstény
túlélését a sebesüléssel (és
a fertõzés veszélyével).
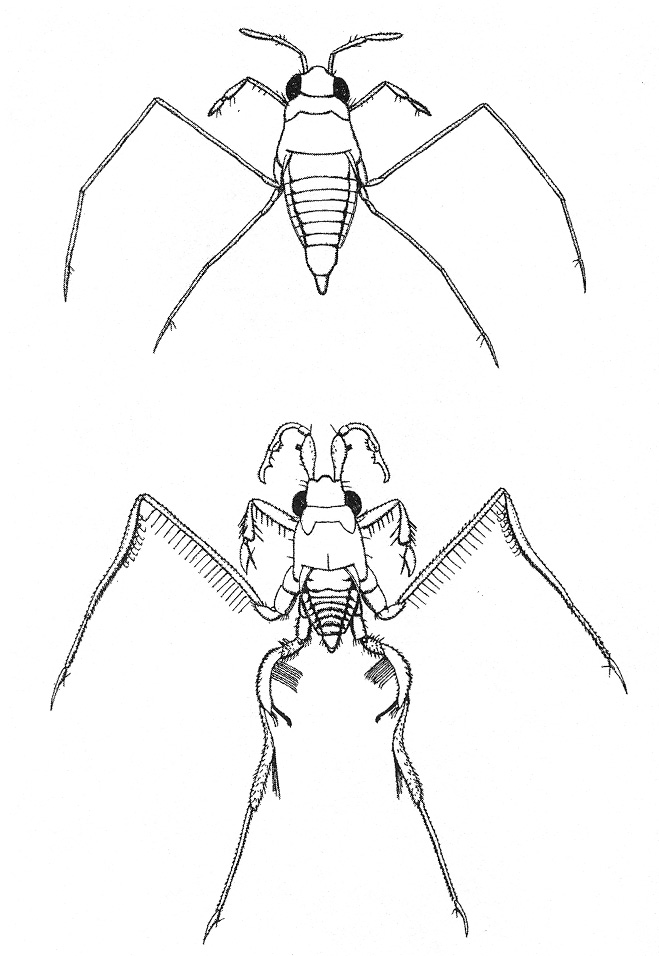
Scatophaga stercoraria trágyalegyek a dulakodás során a nõstényt (kakiba petéz) a trágya alá is nyomhatják, meg is fullad szegény. Gerromorpha vízi molnárpoloskák a kényszerített kopulációkkal gyakran megsebesítik a lányokat. Sok faj hímje plusz függelékeket visel a lábain és csápjain, amivel a lányokat hatékonyan meg lehet ragadni és hiába ficánkol. Számos faj (fõleg rágcsálóknál)
hímje alkalmaz ún. "kopulációs dugót,
ami a nõstény ivari traktusát a kopuláció
után idõlegesen lezárva megnehezíti a
többi hím sikeres közeledését, de a
nõstényt is akadályozza a szabad hölgyválaszban.
|
{kind=link}
{kind=link}
Hím szempontból a nõstények többszörös
párzása mindig hátrányos, mivel csökkenti
az apaság esélyét. A szexuális szelekció
kedvezhet azoknak a hím jellegeknek, melyek növelik a nõstény
újrapárzásának költségeit, kockázatossá
téve azt, és csökkentve az újrapárzás
esélyét. A nõstény genitáliák károsítása,
nõstény fitnessének csökkentése, agresszív
viselkedés, vagy
kényszerített kopuláció mind jelenthet elõnyt
a hím számára. Hím jelleg befolyásolja
a nõstény viselkedést, fogékonyságát,
hajlandóságát.
|
Johnstone és Keller modellje (2000): 1. Intenzív versengés
kialakíthat toxikus mechanizmusokat (mint a muslica-glikoprotein). |
| Drosophila fajok ondófolyadékában találhatóak olyan szekrétumok (pl. az Acp36DE glikoprotein), amelyek a nõstény fogékonyságát csökkenti a késõbbiekben, a többi hím spermáját is bántja, és elõsegíti a tojásrakás megindulását. Emellett valamilyen módon toxikus a nõsténynek, mert az élettartamát csökkenti. Egy kísérletbõl kiderült,
hogy a monogámiára kényszerített (egy
nõsténnyel együtt tartott) hímeknél
34 generáció után kevesebb káros hatással
járt a velük párzó nõstényekre,
azok túlélése és fekunditása nagyobb
volt, mint a monogámiára nem kényszerített
hímekkel (ahol több hímmel is párosodhatnak)
való egyszeri párzás után. A monogámiában
tartott nõstények a nem monogám hímekhez
berakva kevésbé éltek túl, mint a nem
monogámiában tartott lányok, ami azt sugallja,
hogy a promiszkuitás során valamiféle rezisztencia
van fenntartva a fokozott hím-nyomás ellen, ami a monogám
nõstényeknél eltûnik, mert nincs rá
szükség. 47 nemzedék után a monogám
populációk nettó szaporodási sikere nagyobb
volt, mint a versengést megengedõké. A szexuális
szelekció tehát nem minden esetben növeli a fitnesst,
hiszen a káros hatások dominálhatnak. |
{kind=link}
A nõstények a káros hatásokat egyensúlyozandó, szintén mutathatnak morfológiai, kémiai vagy viselkedési adaptációt, kifejleszthetnek olyan rezisztanciajellegeket, mint a hölgyválasz vagy a monogámia, ezekkel mérsékelve a hím- hím versengés okozta nõi költségeket, mint pl. a spermakompetíció miatti spermatoxicitás vagy a ragadozásnak való kitettség, esetleg az udvarlás és erõszakos párzás miatti energetiakai költségek és sérülési veszélyek. Tehát a szexuális szelekció erõssége is alakíthatja a konfliktushoz való adaptálódást.
| Rablólegyeknél
a nemi erõszak áldozatául esõ nõstény
halottnak tetteti magát. |
A nõstények adaptációjára válaszul a hímek újabb alkalmazkodással válaszolhatnak, így állandó szelekciós nyomást gyakorolva egymásra, mint egy fegyverkezési verseny résztvevõi.
![]()
